Dimorphic basidiomycetes, an overview
J.P. Sampaio and A. Fonseca
![]() Dimorphic fungi - what are they?
Dimorphic fungi - what are they?
![]() Yeasts - definitions and diversity
Yeasts - definitions and diversity
![]() Typical basidiomycetous yeasts vs. the yeast stages of Heterobasidiomycetes
Typical basidiomycetous yeasts vs. the yeast stages of Heterobasidiomycetes
![]() Dimorphic fungi - what are they?
Dimorphic fungi - what are they?
The true fungi or Eumycota are currently classified in four phyla: Chytridiomycota, Zygomycota, Ascomycota and Basidiomycota (Alexopoulos et al., 1996; Berbee and Taylor, 1992; Blackwell et al., 1996; Bruns et al., 1991; Taylor, 1995). The typical vegetative form of the Eumycota is an apically elongating walled filament called hypha. This structure is less complex in the Chytridiomycota. Hyphae of the Zygomycota are normally wide and lack septa. Conversely, the hyphae of the Ascomycota and Basidiomycota are narrower, better adapted to thrive in environments with reduced moisture levels and
possess septa at regular intervals.
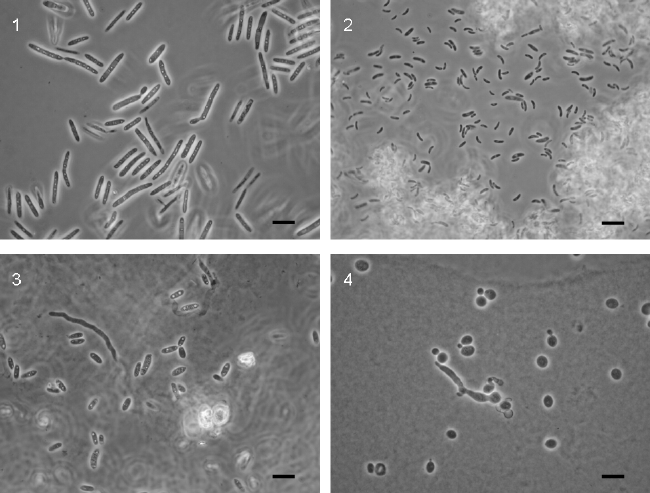
Some fungi belonging to the Ascomycota and Basidiomycota (and also a few members of the Zygomycota) have a distinct vegetative form: the yeast cell. The unicellular growth may result from the division, by budding, of a meiospore (ascospore or basidiospore). In some species, mostly ascomycetes, the entire life cycle takes place in the unicellular stage, as in the case of Saccharomyces cerevisiae Meyen ex Hansen. Other fungi are called dimorphic sensu Oberwinkler (1987), Bandoni (1995), Boekhout et al. (1998), because typically their life cycle has two distinct stages, one unicellular and the other filamentous. In sexual species, the yeast phase results from the budding of meiospores and ends when two sexually compatible yeast cells conjugate and give rise to an usually dikaryotic mycelium, which later will form the sexual structures. Most sexual members of the Basidiomycota having a unicellular stage display also this type of life cycle. Some are plant parasites, like Ustilago maydis (De Candolle) Corda (Fig. 1), others are mycoparasites like Colacogloea peniophorae (Bourd. & Galz.) Oberwinkler & Bandoni (Fig. 2), and others appear to be exclusivelly saprobic like Rhodosporidium toruloides Banno (Fig. 3) or Kondoa malvinella (Fell & Hunter) Yamada, Nakagawa & Banno (Fig. 4).
![]() Figures 1 - 4.
Figures 1 - 4.
As shown in Fig. 5, the realm of dimorphic basidiomycetes is extremely diverse and is distributed among the proposed three classes of the Basidiomycota: Urediniomycetes, Ustilaginomycetes and Hymenomycetes (Swann and Taylor 1993, 1995a, b,c).
![]() Figures 5A - B.
Figures 5A - B.
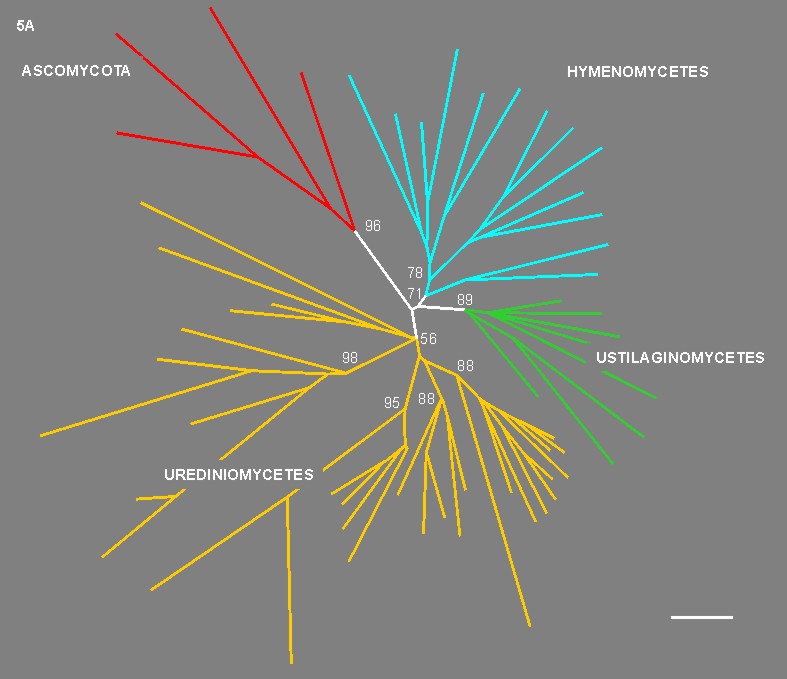
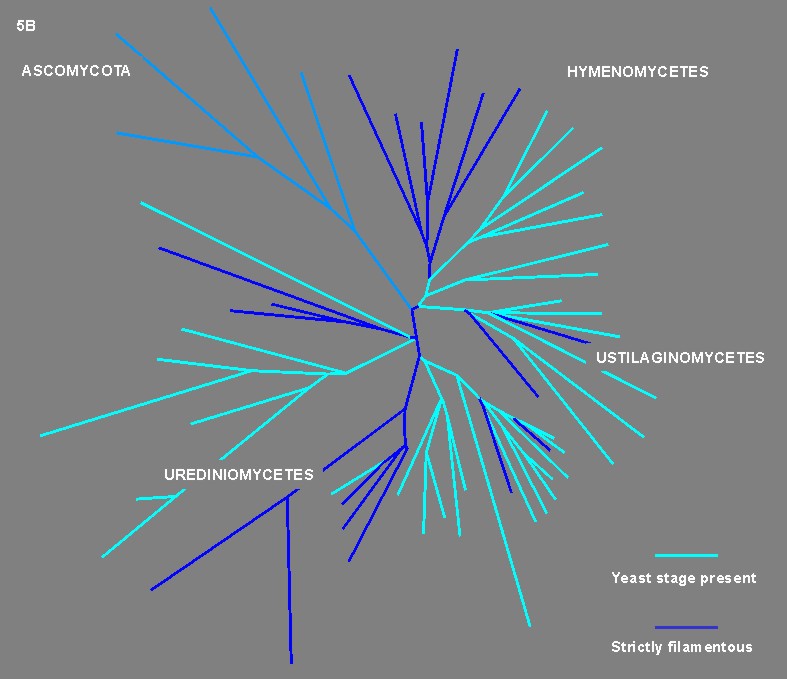
Dimorphic taxa like Microbotryum Léveillé emend. Deml & Oberwinkler, Rhodosporidium Banno, Colacogloea Oberwinkler & Bandoni, Chionosphaera Cox, Agaricostilbum Wright and Occultifur Oberwinkler are classified in the class Urediniomycetes. However, important groups of this class like the rusts (order Uredinales) seem to lack a yeast stage. Among the class Ustilaginomycetes, which includes a vast assemblage of plant parasites, many taxa like Ustilago (Persoon) Roussel, Microstroma Niessl, and Exobasidium Woronin possess a unicellular stage. In the class Hymenomycetes dimorphic taxa are prevalent in the sub-class Tremellomycetidae whereas the sub-class Hymenomycetidae includes only filamentous taxa. Examples of members of the Tremellomycetidae are Tremella
Persoon, Filobasidium Olive and Filobasidiella Kwon-Chung. Since many taxa exhibiting ancestral traits (e.g. simple septal pores) are dimorphic and the occurrence of a dimorphic state in the Basidiomycota is much more widespread than the dissemination of strictly filamentous taxa, it can be hypothesized that the ancestral basidiomycetous lineage was dimorphic. The dimorphic basidiomycetes exhibit some common traits, as follows:
![]() The sexual stage develops in the filamentous phase (exception: Xanthophyllomyces Golubev)
The sexual stage develops in the filamentous phase (exception: Xanthophyllomyces Golubev)
![]() The haploid stage corresponds to the unicellular stage
The haploid stage corresponds to the unicellular stage
![]() The unicellular stage results from basidiospore germination
The unicellular stage results from basidiospore germination
![]() When parasitic, the parasitic stage corresponds to the filamentous (and sexual) stage and the unicellular stage corresponds to a saprobic phase, probably related to dispersal
When parasitic, the parasitic stage corresponds to the filamentous (and sexual) stage and the unicellular stage corresponds to a saprobic phase, probably related to dispersal
![]() Yeasts - definitions and diversity
Yeasts - definitions and diversity
For historical reasons, mostly due to the relevance of yeasts in the production of alcoholic beverages, the taxonomy of unicellular fungi evolved separately from the other areas of mycology. On one hand yeast cells have few morphological attributes upon which to base a classification, as was the rule during many years for filamentous fungi. On the other hand, discriminating yeast species on the basis of different physiological attributes had an obvious industrial importance.
The term “yeast” has had different meanings for mycologists and “zymologists”. For some, “yeast” was (is) used to designate a taxonomic group. This meaning of “yeast” should be rejected because it is now clear that the yeasts do not form a natural group, i.e. yeasts are clearly polyphyletic. Others have used the terms “yeast” and “yeast-like”, the first for the “true” yeasts (ascomycetous and strictly unicellular) and the second for “deviating” yeasts that in certain cases became filamentous (e.g. dimorphic basidiomycetes) or produced dark pigments (e.g. Aureobasidium spp). The difficulties to define “yeast-like” are obvious and this concept varied from one author to the other. Finally, there is the ontological concept of “yeast”, according to which “yeast” simply describes a particular state in the life cycle of a fungus (Flegel, 1977). In view of this concept, the list of yeasts should be enlarged to include organisms like Symbiotaphrina Kühlwein & Jurzitza ex Gams & von Arx (Euascomycetes), or Taphrina Fr. (Archiascomycetes), as well as all the dimorphic basidiomycetes regardless of their ecological strategies and presence or absence of a fruitbody.
Yeast taxonomists have recognized certain dimorphic taxa as belonging to the realm of basidiomycetous yeasts whereas other taxa were overlooked because they were parasitic or produced a fruitbody. The former were isolated in pure cultures, which were then characterized in the laboratory with respect to physiological and biochemical characteristics and also with respect to certain morphological traits. The latter were found after laborious field work, by collection of samples of fruitbodies or of infected plants or other fungi which were then investigated under the microscope. Dried material was preserved for reference but the isolation of living cultures was not considered essential. Three factors might explain this: (a) lack of knowledge of how
to cultivate the microorganisms and facilities to achieve it; (b) lack of motivation to obtain living cultures since normally they exhibited only the saprobic yeast stage and not the basidial stage; (c) difficulty to distinguish between the correct culture and other yeast contaminants present in the original sample.
Moreover yeast taxonomists have always paid attention to asexual stages and much effort was made to integrate them in the classification system. In fact, most taxa traditionally recognized as typical basidiomycetous yeasts are asexual, whereas most of the organisms traditionally studied by mycologists and labelled as dimorphic heterobasidiomycetes are sexual.
Molecular methods, especially DNA sequence analyses, have demonstrated, in many cases, that the taxa described by mycologists and the taxa described by “zymologists” were phylogenetically related. For example, a close relative of the smut fungus Ustilago is the anamorphic genus Pseudozyma Bandoni emend. Boekhout. Whereas for some Pseudozyma species a teleomorphic connection with Ustilago has been established, for must species in Pseudozyma no such relationship has been demonstrated. Does Pseudozyma represent the anamorphic and saprobic state of Ustilago or is it formed by Ustilago-like organisms that have lost their phytoparasitic (and sexual) capacity?
The Ustilago / Pseudozyma example, and several other cases like the relationship of Colacogloea, Microbotryum and Microstroma with different species of Rhodotorula, illustrate the need for an integrated approach in order to obtain a unified classification system of dimorphic basidiomycetes.
![]() About this page
About this page
Sampaio, J.P. and Fonseca, A. 2002. Dimorphic Basidiomycetes, an overview. Dimorphic Basidiomycetes WWW Project.
Version 01, saved 20 June 2002.
José Paulo Sampaio: jss@mail.fct.unl.pt
Álvaro Fonseca: amrf@mail.fct.unl.pt
Centro de Recursos Microbiológicos, Secção Autónoma de Biotecnologia, Faculdade de Ciências e Tecnologia, Universidade Nova de Lisboa, 2829-516 Caparica, Portugal.
Page copyright © 2002 J.P. Sampaio and A. Fonseca
Dimorphic Basidiomycetes design and icons copyright © 2002 J.P. Sampaio