The classification of dimorphic basidiomycetes
J.P. Sampaio and R. Bauer
![]() Class Hymenomycetes (Tremellomycetidae)
Class Hymenomycetes (Tremellomycetidae)
The current tripartite classification concept of the Basidiomycota is mainly based on sequence analyses of 18S rRNA (Taylor, 1995; Swann and Taylor, 1993; 1995a; 1995b; 1995c). As a consequence of those studies, the classes Urediniomycetes, Ustilaginomycetes and Hymenomycetes have been proposed (Swann and Taylor 1995a). Analysis of the D1/D2 region of the 26S rDNA shows basically the same pattern (Begerow et al., 1997). The data derived from the analysis of cell wall carbohydrates correlate well with the molecular phylogenies (Prillinger et al., 1990; 1991a; 1991b; 1993) and the same holds for the structure of the septal pores and the shape of the spindle pole bodies (McLaughlin et al., 1995a; 1995b and references therein).
This class, as proposed by Swann and Taylor (1995a, 1995b) is characterized by simple septal pores without membrane caps and disc-like spindle pole bodies (Bauer and Oberwinkler 1994, Wells 1994, McLaughlin et al., 1995b). Except for a few species, the basidia are transversally septate. Mannose is the major cell wall carbohydrate, glucose, fucose and rhamnose are the less prevalent neutral sugars and xylose is not present (Prillinger et al., 1991b; 1993).
Within the Urediniomycetes, Swann et al. (1999) defined the subclass Microbotryomycetidae Swann for a monophyletic group (based on 18S rRNA gene sequences) encompassing teleomorphic genera such as Microbotryum Léveillé emend. Vánky, Rhodosporidium Banno, Leucosporidium Fell, Statzell, Hunter & Phaff, Colacogloea Oberwinkler & Bandoni, Kriegeria Bres. and Heterogastridium Oberwinkler & Bauer. This subclass comprises predominantly real or potential mycoparasites having colacosomes (Bauer and Oberwinkler,1991a) and the urediniomycetous smut fungi (Bauer et al.,1997). Fell et al. (2000; 2001) and Scorzetti et al. (2002) have analysed the D1/D2 region of the 26S rDNA of a vast number of (typical) basidiomycetous yeast species and their phylogenetic trees depict a considerable number of anamorphic species of the genera Rhodotorula Harrison and Sporobolomyces Kluyver & van Niel as belonging to the same monophylum. Recently, Sampaio et al. (2003) proposed the order Leucosporidiales Sampaio, Weiss & Bauer for Leucosporidium scottii and closely related sexual and asexual species (Sampaio et al. 2003). In the same report the order Sporidiobolales Sampaio, Weiss & Bauer was erected for Rhodosporidium, Sporidiobolus Nyland and allied asexual taxa.
In 2001, Swann et al. treated most of the remaining members of the class Urediniomycetes in two additional subclasses, Agaricostilbomycetidae and Urediniomycetidae. Whereas the known members of the Agaricostilbomycetidae are dimorphic, most of the taxa belonging to the Urediniomycetidae are filamentous. Examples of teleomorphic taxa of the Agaricostilbomycetidae are Agaricostilbum Wright, a fungus associated with palms, Chionosphaera Cox, which produces holobasidia, and Kondoa Yamada, Nakagawa & Banno emend. Fonseca, Sampaio & Fell, a genus investigated by Fonseca et al. (2000) who found that it lacks teliospores but forms transversally septate basidia which are able to produce forcibly discharged basidiospores. Based on sequence analyses, the mitosporic genera Kurtzmanomyces Yamada, Itoh, Kawasaki, Banno & Nakase emend. Sampaio, Sterigmatomyces Fell emend. Yamada & Banno and most of the species of Bensingtonia Ingold emend. Nakase & Boekhout belong to the Agaricostilbomycetidae (Fell et al., 2000; 2001; Fonseca et al., 2000; Hamamoto and Nakase, 2000; Scorzetti et al., 2002).
The subclass Urediniomycetidae is mainly composed of plant parasites. The most relevant group of this subclass, in terms of numbers of species and impact in human activities, are the rusts (order Uredinales). Interestingly, taxa with yeast stages are almost absent in the Urediniomycetidae since budding cells are currently known only in species of the genus Septobasidium Pat.
The classification of the Urediniomycetes is far from being settled due, among other factors, to the lack of detailed information for many species. Besides the three subclasses mentioned above, other groups have been detected based on ultrastructural and molecular studies.
One of the unnamed clades includes the sexual genera Naohidea Oberwinkler, Occultifur Oberwinkler, Sakaguchia Yamada, Maeda & Mikata and several asexual species of the genera Rhodotorula and Sporobolomyces. The genera Erythrobasidium Hamamoto, Sugiyama & Komagata and Bannoa Hamamoto belong also to this clade. Erythrobasidium and Bannoa were originally described for sexual species (Hamamoto et al., 1988; 2002) but the postulated basidia of Erytrhobasidium were later interpreted as conidiogenic structures (Sampaio et al., 1999). Bannoa is morphologically and phylogenetically close to Erythrobasidium and also lacks true basidia (Sampaio unpubl.). Takashima et al. (2000) determined the carbohydrate composition for representative taxa of this clade and observed in all cases the absence of fucose, thus providing an additional criterion for the separation of this group in a taxon of its own.
Another distinct clade corresponds to the order Atractiellales sensu Oberwinkler and Bauer (1989). The taxa in this order do not have a yeast stage and belong to the genera Atractiella Sacc., Helicogloea Pat., Phleogena Link and Saccoblastia Möller. Molecular studies are scarce for this group but relevant ultrastructural markers like the type of septal pore and the presence of symplechosomes (Oberwinkler and Bauer, 1989, Bauer and Oberwinkler, 1991b) - designated as microscala by McLaughlin (1990) - indicate that those four genera share a close common ancestor.
Recently, Bauer et al. (2003) proposed the order Classiculales Bauer, Begerow, Oberwinkler & Marvanová for urediniomycetous taxa having the uredinalian type of septal pores in combination with tremelloid haustorial cells.
Below, we present an annotated outline of the class Urediniomycetes which attempts to integrate data from several authors, namely Bandoni, Bauer, Fell, Oberwinkler, Swann and Vánky.
CLASS UREDINIOMYCETES
SUB-CLASS MICROBOTRYOMYCETIDAE
Taxa having colacosomes and taxa related to them
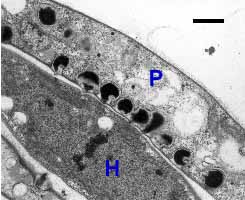
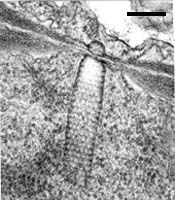
Colacosomes of Colacogloea peniophorae. Colacosomes are the dark electron-dense structures in P (hypha of the parasite); H, hypha of Hyphoderma praetermissum (host). Bar = 1 µm; © R. Bauer.
Order Cryptomycocolacales Oberwinkler & Bauer
Family Cryptomycocolacaceae Oberwinkler & Bauer
Cryptomycocolax Oberwinkler & Bauer
Colacosiphon Kirschner, Bauer & Oberwinkler
Order Heterogastridiales Oberwinkler & Bauer
Family Heterogastridiaceae Oberwinkler & Bauer
Heterogastridium Oberwinkler & Bauer
Order Leucosporidiales Sampaio, Weiss & Bauer
Leucosporidium Fell, Statzell, Hunter & Phaff
Leucosporidiella Sampaio
Mitosporic
Mastigobasidium Golubev
Order Sporidiobolales Sampaio, Weiss & Bauer
Family Sporidiobolaceae Moore emend. Sampaio, Weiss & Bauer
Sporidiobolus Nyland
Sporobolomyces Kluyver & van Niel
Mitosporic, not monophyletic
Rhodosporidium Banno
Rhodotorula Harrison
Mitosporic, not monophyletic
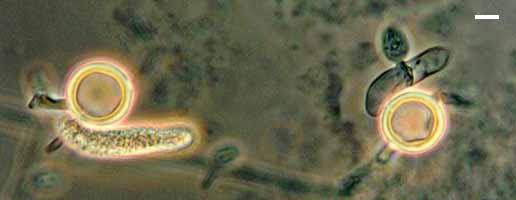
Teliospores, basidia and basidiospores of Sporidiobolus salmonicolor. Bar = 5 µm; © J.P. Sampaio.
Order Microbotryales Bauer & Oberwinkler
Phytoparasitic members of the Basidiomycota having transversely septate basidia with multiple production of sessile basidiospores and only intercellular hyphae (Bauer et al., 1997).
Family Microbotryaceae MooreMembers of the Microbotryales having poreless septa at maturity (Bauer et al., 1997).
Bauerago Vánky
Liroa Ciferri
Microbotryum Léveillé emend. Vánky
Approximately 75 species parasitizing dicotyledonous plants
Sphacelotheca de Bary emend. Langdon et Fullerton
Zundeliomyces Vánky
Family Ustilentylomataceae Bauer & Oberwinkler
Members of the Microbotryales having simple septal pores (Bauer et al., 1997).
Aurantiosporium Piepenbring, Vánky & Oberwinkler
Fulvisporium Vánky
Ustilentyloma Savile
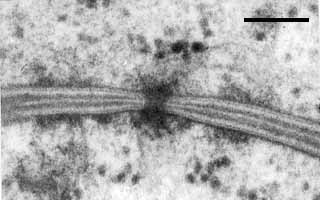
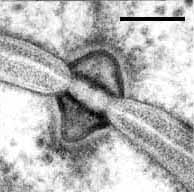
Septal pore of Ustilentyloma fluitans. Bar = 0.1 µm; © R. Bauer.
Taxa of the Microbotryomycetidae not ascribed to any order
Family Krieglsteineraceae Pouzar
Krieglsteinera Pouzar
Family Camptobasidiaceae Moore
Camptobasidium Marvanová & Suberkropp
Atractocolax Kirschner, Bauer & Oberwinkler
Bensingtonia Ingold emend. Nakase & Boekhout
Mitosporic, not monophyletic
Colacogloea Oberwinkler & Bandoni
Kriegeria Bres.
SUB-CLASS AGARICOSTILBOMYCETIDAE
Order Agaricostilbales Oberwinkler & Bauer
Family Agaricostilbaceae Oberwinkler & Bauer
Agaricostilbum Wright emend. Wright, Bandoni & Oberwinkler
Family Chionosphaeraceae Oberwinkler & Bandoni
Chionosphaera Cox
Stilbum Tode ex Mérat
Fibulostilbum Seifert & Oberwinkler
Genera of the Agaricostilbomycetidae not ascribed to any order
Bensingtonia Ingold emend. Nakase & Boekhout
Mitosporic, not monophyletic
Kondoa Yamada, Nakagawa & Banno emend. Fonseca, Sampaio & Fell
Kurtzmanomyces Yamada, Itoh, Kawasaki, Banno & Nakase emend. Sampaio
Mitosporic
Mycogloea Olive
Sporobolomyces Kluyver & van Niel
Mitosporic, not monophyletic
Sterigmatomyces Fell emend. Yamada & Banno
Mitosporic
Spiculogloea Roberts
Zygogloea Roberts
SUB-CLASS UREDINIOMYCETIDAE
Order Uredinales Arthur
14 families, 164 genera, aprox. 7000 species
Order Septobasidiales Couch ex Donk
Family Septobasidiaceae Maire
Auriculoscypha Reid and Manimohan
Coccidiodictyon Oberwinkler
Ordonia Racib.
Septobasidium Pat.
Uredinella Couch
Family Pachnocybaceae Oberwinkler & Bauer
Pachnocybe Berk.
Order Platygloeales Moore
Platygloea Schroeter sensu stricto (P. disciformis and related species)
Taxa of the Urediniomycetidae not ascribed to any order
Family Eocronartiaceae Jülich
Eocronartium Atkinson
Helicobasidium Pat.
Herpobasidium Lind emend. Oberw & Bandoni
Insolibasidium Oberwinkler & Bandoni
Jola Möller
Platycarpa Couch emend. Oberwinkler & Bandoni
Ptechetelium Oberwinkler & Bandoni
Groups of Urediniomycetes for which higher rank classification is incomplete
Order Atractiellales Oberwinkler & Bandoni
Family Hoehnelomycetaceae Jülich
Atractiella Sacc.
Family Phleogenaceae Weese
Phleogena Link
Family Helicogloeaceae Jülich
Helicogloea Pat.
Saccoblastia Möller
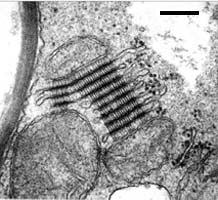
Symplechosomes of Saccoblastia farinacea. Bar = 0.25 µm; © R. Bauer.
Order Classiculales Bauer, Begerow, Oberwinkler & Marvanová
Family Classiculaceae Bauer, Begerow, Oberwinkler & Marvanová
Classicula Bauer, Begerow, Oberwinkler & Marvanová
Jaculispora H. J. Huds. & Ingold
Mitosporic
Order Cystobasidiales (Bauer & Sampaio, in prep.)
Septal pore and cystosome of Occultifur internus. Bar = 0.2 µm; © R. Bauer.
Bannoa Hamamoto
Cystobasidium (Lagerh.) Neuhoff
Naohidea Oberwinkler
Occultifur Oberwinkler
Sakaguchia Yamada, Maeda & Mikata
Erythrobasidium Hamamoto, Sugiyama & Komagata
Rhodotorula Harrison
Mitosporic, not monophyletic
Sporobolomyces Kluyver & van Niel
Mitosporic, not monophyletic
Family Mixiaceae Kramer
Mixia Kramer
The Ustilaginomycetes are predominantly plant-parasites and include ca. 1400 species. They can be distinguished from the other basidiomycetes because they have a distinctive cell wall carbohydrate composition with dominance of glucose and absence of xylose (Prillinger et al.,1990; 1993). The type B secondary structure of the 5S rRNA is shared with the Hymenomycetes (Gottschalk and Blanz, 1985) and, like the Urediniomycetes, they lack parenthesomes at the septal pores (Bauer et al., 1997).
Several recent publications have dealt with the classification of the Ustilaginomycetes (Bauer et al., 1997; Bauer et al., 2001; Begerow et al., 1997) and a comprehensive classification scheme based mainly on the characteristics of host-parasite interactions and septal pore apparatus has been proposed. In general, sequence data gives support to the classification proposals listed below, derived mainly from Bauer et al. (2001) and Begerow et al. (2002). The supra-generic classification of mitosporic taxa belonging to the Ustilaginomycetes is also being improved. The order Malasseziales Moore emend. Begerow, Bauer & Boekhout was redefined and presently accommodates the species of Malassezia Baillon, a genus of the Exobasidiomycetidae (Begerow et al., 1997). The genus Pseudozyma Bandoni emend. Boekhout seems to represent the asexual counterpart of Ustilago (Persoon) Roussel, whereas Tilletiopsis Derx ex Derx belongs to the Exobasidiomycetidae but is polyphyletic (Begerow et al., 2000). Sympodiomycopsis paphiopedili Sugiyama, Tokuoka & Komagata and three species of Rhodotorula, namely Rh. bacarum (Buhagiar) Rodrigues de Miranda & Weijman, Rh. hinnulea (Shivas & Rodrigues de Miranda) Rodrigues de Miranda & Weijman and Rh. phylloplana (Shivas & Rodrigues de Miranda) Rodrigues de Miranda & Weijman belong to the order Microstromatales of the Exobasidiomycetidae (Fell et al., 2001; Begerow et al., 2001). The classification system of the Ustilaginomycetes is presented below.
CLASS USTILAGINOMYCETES Bauer, Oberwinkler & Vánky
SUB-CLASS ENTORRHIZOMYCETIDAE Bauer & Oberwinkler
Members of the Ustilaginomycetes having local interaction zones and pores without membranous bands or caps (Bauer et al., 1997).
Order Entorrhizales Bauer & Oberwinkler
Family Entorrhizaceae Bauer & Oberwinkler
Entorrhiza Weber
SUB-CLASS USTILAGINOMYCETIDAE Jülich emend. Bauer & Oberwinkler
Members of the Ustilaginomycetes having enlarged interaction zones. Septa have pores with membranous caps or they are poreless at maturity.
Order Urocystales Bauer & Oberwinkler
Family Melanotaeniaceae Begerow, Bauer & Oberwinkler
Exoteliospora Bauer, Oberwinkler & Vánky
Melanotaenium de Bary
Yelsemia Walker
Family Doassansiopsaceae Begerow, Bauer & Oberwinkler
Doassansiopsis (Setchell) Dietel
Family Urocystaceae Begerow, Bauer & Oberwinkler
Mundkurella Thirumalachar
Urocystis Rabenhorst ex Fuckel
Ustacystis Zundel
Vankya Ershad
Septal pore of Ustacystis waldsteiniae. Bar = 0.1 µm; © R. Bauer.
Order Ustilaginales Clinton emend. Bauer & Oberwinkler
Family Ustilaginaceae L. Tulasne & C. Tulasne emend. Bauer & Oberwinkler
Anthracoidea Brefeld
Cintractia Cornu
Cintractiella K. B. Boedijn
Clintamra Cordas & Durán
Dermatosorus Sawada ex Ling
Farysia Raciborski
Farysporium Vánky
Franzpetrakia Thirumalachar & Pavgi emend. Guo, Vánky & Mordue
Geminago Vánky et Bauer
Heterotolyposporium Vánky
Kuntzeomyces Hennings ex Saccardo & Sydow
Leucocintractia Piepenbring, Begerow & Oberwinkler
Macalpinomyces Langdon & Fullerton emend. Vánky
Melanopsichium Beck
Moesziomyces Vánky
Moreaua Liou & Cheng
Orphanomyces Saville
Pericladium Passerini emend. Mundkur
Planetella Saville
Pseudozyma Bandoni emend. Boekhout
Mitosporic "Rhodotorula" acheniorum (Buhagiar & Barnett) Rodrigues de MirandaMitosporic
Schizonella Schröter
Sporisorium Ehrenberg. ex Link
Stegocintractia Piepenbring, Begerow & Oberwinkler
Testicularia Klotzsch
Tolyposporium Woronin ex Schröter
Tranzscheliella Lavrov
Trichocintractia Piepenbring, Begerow & Oberwinkler
Uleiella Schröter
Ustanciosporium Vánky emend. Piepenbring
Ustilago (Persoon) Roussel
Websdanea Vánky
Family Glomosporiaceae Cifferi emend. Begerow, Bauer & Oberwinkler
Glomosporium Kochman
Thecaphora Fingerhuth emend. Vánky
Tothiella Vánky
Family Mycosyringaceae Bauer & Oberwinkler
Mycosyrinx Beck
SUB-CLASS EXOBASIDIOMYCETIDAE Jülich emend. Bauer & Oberwinkler
Members of the Ustilaginomycetes having local interaction zones. Septa have pores with membranous bands or caps or they are poreless at maturity.
Order Malasseziales Moore emend. Begerow, Bauer & Boekhout
Malassezia Baillon
Order Georgefischeriales Bauer, Begerow & Oberwinkler
Family Georgefischeriaceae Bauer, Begerow & Oberwinkler
Georgefischeria Thirumalachar & Narasimhan emend Gandhe
Jamesdicksonia Thirumalachar, Pavgi & Payak
Family Tilletiariaceae Moore
Phragmotaenium Bauer, Begerow, Nagler & Oberwinkler
Tilletiaria Bandoni & Johri
Tilletiopsis flava (Tubaki) Boekhout
Mitosporic
Tilletiopsis fulvescens Gokhale
Mitosporic
Tolyposporella Atkinson
Family Eballistraceae (Bauer, Begerow, Nagler & Oberwinkler)
Eballistra Bauer, Begerow, Nagler & Oberwinkler
Order Tilletiales Kreisel ex Bauer & Oberwinkler
Family Tilletiaceae L & C Tullasne emend. Bauer et Oberwinker
Conidiosporomyces Vánky
Erratomyces Piepenbring & Bauer
Ingoldiomyces Vánky
Neovossia Körnicke
Oberwinkleria Vánky & Bauer
Tilletia L. & C Tullasne
Order Microstromatales Bauer & Oberwinkler
Family Microstromataceae Jülich
"Cerinosterus" cyanescens (de Hoog & de Vries) Moore
Microstroma Niessl
"Rhodotorula" bacarum (Buhagiar) Rodrigues de Miranda & Weijman
Mitosporic
"Rhodotorula" phylloplana (Shivas & Rodrigues de Miranda) Rodrigues de Miranda & WeijmanMitosporic
Sympodiomycopsis Sugiyama, Tokuoka & Komagata
Mitosporic
Family Volvocisporaceae Begerow, Bauer & Oberwinkler
Volvocisporium Begerow, Bauer & Oberwinkler
Superorder Exobasidianae Bauer & Oberwinkler
Members of the Exobasididiomycetidae having simple pores and interaction apparatus
Order Entylomatales Bauer & Oberwinkler
Family Entylomataceae Bauer & Oberwinkler
Entyloma de Bary
Order Doassansiales Bauer & Oberwinkler
Family Melaniellaceae Bauer, Vánky, Begerow & Oberwinkler
Mellaniella Bauer, Vánky, Begerow & Oberwinkler
Family Doassansiaceae (Azb. & Karat.) Moore emend. Bauer & Oberwinkler
Burillia Setchell
Doassansia Cornu
Doassinga Vánky, Bauer & Begerow
Heterodoassansia Vánky
Nannfeldtiomyces Vánky
Narasimhania Thirumalachar et Pavgi emend. Vánky
Pseudodermatosorus Vánky
Pseudodoassansia (Setchell) Vánky
Pseudotracya Vánky
Tracya H. & P. Sydow
Family Rhamphosporaceae Bauer & Oberwinkler
Rhamphospora Cunningham
Order Exobasidiales Hennings emend. Bauer & Oberwinkler
Family Brachybasidiaceae Gäumann
Brachybasidium Gäumann
Dicellomyces Olive
Exobasidiellum Donk
Kordyana Racib.
Proliferobasidium Cunningham
Family Exobasidiaceae Hennings
Exobasidium Woronin
Muribasidiospora Kamat e & Rajendren
Family Cryptobasidiaceae Malençon ex Donk
Botryoconis H. & P. Sydow
Clinoconidium Pat.
Coniodictyum Hariot & Pat.
Drepanoconis Schröter & P. Hennings
Laurobasidium Jülich
Family Graphiolaceae Fischer
Graphiola Poiteau
Stylina H. Sydow
Taxa of the Exobasidiomycetidae not ascribed to any family
Ceraceosorus bombacis (Bakshi) Bakshi
Tilletiopsis albescens Gokhale
Mitosporic
Tilletiopsis cremea Tubaki
Mitosporic
Tilletiopsis lilacina Tubaki
Mitosporic
Tilletiopsis minor Nyland
Mitosporic
Tilletiopsis pallescens Gokhale
Mitosporic
Tilletiopsis washingtonensis Nyland
Mitosporic
![]() Class Hymenomycetes (Tremellomycetidae)
Class Hymenomycetes (Tremellomycetidae)
The class Hymenomycetes as defined by Swann and Taylor (1995a; 1995b; 1995c) includes the complex-septate basidiomycetes (i.e. with dolipore), including the gasteromycetes. Moreover, the members of the Hymenomycetes have glucose as the major cell wall carbohydrate component and, at variance with the Urediniomycetes and Ustilaginomycetes, xylose is present. Swann and Taylor (1995b) distinguished two subclasses, viz. Hymenomycetidae and Tremellomycetidae. A few traits other than DNA sequences differentiate the two subclasses. Sacculate membrane elements are normally observed at the vicinity of the dolipore of the Tremellomycetidae (in some cases they are lacking), whereas the members of the Hymenomycetidae have smooth membranous septal pore caps of various types, but not sacculate. In addition, whereas the vast majority of the Tremellomycetidae produces a yeast phase, such ontogenetic stage is not observed in the Hymenomycetidae. Therefore, the present review will deal only with the Tremellomycetidae. The classification system presented below is based mainly on the revisions of Wells (1994) and Wells and Bandoni (2001) which are derived mostly from comparisons of morphological and ultrastructural data. New teleomorphic genera described in Kirschner et al. (2001) and Sampaio et al. (2002) are also included. Although most mitosporic species of the Tremellomycetidae have been investigated in comprehensive sequence analysis studies (Fell et al., 2000; 2001; Scorzetii et al., 2002), several sexual taxa have not yet been studied using the same approaches. It is anticipated that relevant systematic changes will be proposed when such species are investigated by molecular phylogenetic methods.
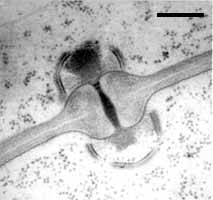
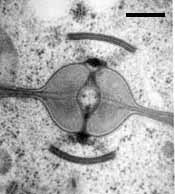
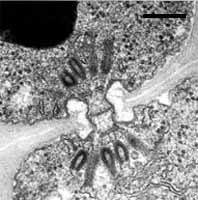
Septal pore types in the Hymenomycetes.
Left: Dolipore with perfurated parenthesomes of Schizophyllum commune (Bar = 0.25 µm; © R. Bauer).
Center: Dolipore with continuous parenthesomes of Tulasnella sp. (Bar = 0.25 µm; © R. Bauer).
Left: Dolipore with cup-shaped parenthesomes of Tremella sp. (Bar = 0.2 µm; © R. Bauer).
CLASS HYMENOMYCETES
SUB-CLASS TREMELLOMYCETIDAE
Order Cystofilobasidiales Boekhout & Fell
Itersonilia Derx
"Cryptococcus" aquaticus (Jones & Slooff) Rodrigues de Miranda & Weijman
"Cryptococcus" huempii (Ramírez & González) Roeijmans, van Eijk & Yarrow
"Cryptococcus" macerans (Frederiksen) Phaff & Fell
Cystofilobasidium Oberwinkler & Bandoni
Mrakia Yamada & Komagata
Tausonia Bab’eva
Mitosporic
"Trichosporon" pullulans (Lindner) Diddens & Lodder
Udeniomyces Nakase & Takematsu
Mitosporic
Xanthophyllomyces Golubev
Order Trichosporonales Boekhout & Fell
"Cryptococcus" curvatus (Diddens & Lodder) Golubev
"Cryptococcus" humicola (Daszewska) Golubev
Trichosporon Behrend
Mitosporic
Order Filobasidiales Jülich
Family Filobasidiaceae Olive
Cryptococcus Vuillemin
Mitosporic, not monophyletic
Filobasidium Olive
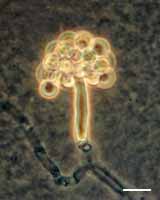
Basidium and basidiospores of Filobasidium floriforme. Bar = 10 µm; © J.P. Sampaio.
Family Carcinomycetaceae Oberwinkler & Bandoni
Carcinomyces Oberwinkler & Bandoni
Christiansenia Hauerslev
Syzygospora Martin
Order Tremellales Rea emend. Bandoni
Family Cuniculitremaceae Sampaio & Kirschner
Cuniculitrema Sampaio & Kirschner
Fellomyces Yamada & I. Banno
Mitosporic
Kockovaella Nakase, Banno & Yamada
Mitosporic
Sterigmatosporidium Kraepelin & Schulze
Mitosporic
Family Phragmoxenidiaceae Oberwinkler & Bauer
Phragmoxenidium Oberwinkler
Phyllogloea Lowy
Family Sirobasidiaceae Möller
Fibulobasidium Bandoni
Sirobasidium Lagerh. & Pat.
Family Tremellaceae Fries emend. Bandoni
Bullera Derx
Mitosporic, not monophyletic
Bulleribasidium Sampaio, Weiss & Bauer
Bulleromyces Boekhout & Fonseca
Cryptococcus Vuillemin
Mitosporic, not monophyletic
Dioszegia Zsolt emend. Takashima, Deak & Nakase
Filobasidiella Kwon-Chung
Holtermannia Sacc. & Traverso
Papiliotrema Sampaio, Weiss & Bauer
Sirotrema Bandoni
Tremella Pers.
Not monophyletic
Tremellina Bandoni
Mitosporic
Trimorphomyces Bandoni & Oberw.
Tsuchiyaea Yamada, Kawasaki, Itoh, Banno & Nakase
Mitosporic
Xenolachne Rogers
Family Tetragoniomycetaceae Oberwinkler & Bandoni
Tetragoniomyces Oberwinkler & Bandoni
Family Rhynchogastremataceae Oberwinkler & Metzler
Rhynchogastrema
Metzler & OberwinklerReferences
Bauer, R. and Oberwinkler, F. 1991a. The colacosomes: new structures at the host-parasite interface of a mycoparasitic basidiomycete. Botanica Acta 104: 53-57.
Bauer, R. and Oberwinkler, F. 1991b. The symplechosome: a unique cell organelle of some basidiomycetes. Botanica Acta 104: 93-97.
Bauer, R. and Oberwinkler, F. 1994. Meiosis, septal pore architecture and systematic position of the heterobasidiomycetous fern parasite Herpobasidium filicinum. Canadian Journal of Botany 72: 1229-1242.
Bauer, R., Oberwinkler, F. and Vánky, K. 1997. Ultrastructural markers and systematics in smut fungi and allied taxa. Canadian Journal of Botany. 75: 1273-1314.
Bauer, R., Begerow, D, Oberwinkler, F. Piepenbring, M. and Berbee, ML. 2001. Ustilaginomycetes In The Mycota – vol VII, Systematics and Evolution, D.J. McLaughlin, E.G. McLaughlin and P.A. Lemke (eds.), Springer-Verlag, Berlin, pp 58-83.
Bauer, R., Begerow, D., Oberwinkler, F. and Marvanová, L. 2003. Classicula: the teleomorph of Naiadella fluitans. Mycologia (in press).
Begerow, D., Bauer, R. and Oberwinkler, F. 1997. Phylogenetic studies on nuclear large subunit ribosomal DNA sequences of smut fungi and related taxa. Canadian Journal Botany 75: 2045-2056.
Begerow, D., Bauer, R. and Boekhout, T. 2000. Phylogenetic placements of ustilaginomycetous anamorphs as deduced from nuclear LSU rDNA sequences. Mycological Research 104: 53-60.
Begerow, D., Bauer, R. and Oberwinkler, F. 2001. Muribasidiospora: Microstromatales or Exobasidiales? Mycological Research 105: 798-810.
Begerow, D., Bauer, R. and Oberwinkler, F. 2002. The Exobasidiales: an evolutionary hypothesis Mycological Progress 1: 187-199.
Fell, J.W., Boekhout, T., Fonseca, A., Scorzetti, G. and Statzell-Tallman, A. 2000. Biodiversity and systematics of basidiomycetous yeasts as determined by large-subunit rDNA D1/D2 domain sequence analysis. International Journal of Systematic and Evolutionary Microbiology 50: 1351-1371.
Fell, J.W., Boekhout, T., Fonseca, A., Sampaio, J.P. 2001. Basidiomycetous yeasts. In The Mycota – vol VII, Systematics and Evolution, D.J. McLaughlin, E.G. McLaughlin and P.A. Lemke (eds.), Springer-Verlag, Berlin, pp 3-35.
Fonseca, A., Sampaio, J.P., Inácio, I. and Fell, J.W. 2000. Emendation of the basidiomycetous yeast genus Kondoa and the description of Kondoa aeria sp. nov. Antonie van Leeuwenhoek 77: 293-302.
Gottschalk, M. and Blanz, P.A. 1985. Untersuchungen an 5S ribosomalen Ribonucleinsäuren als Beitrag zur Klärung von Systematik und Phylogenie der Basidiomyceten. Zeitschrift für Mycologie 51: 205-243.
Hamamoto, M. and Nakase, T. 2000. Phylogenetic analysis of the ballistoconidium-forming yeast genus Sporobolomyces based on 18S rDNA sequences. International Journal of Systematic and Evolutionary Microbiology 50: 1373-1380.
Hamamoto, M., Sugiyama, J. and Komagata, K. 1988. Transfer of Rhodotorula hasegawae to a new basidiomycetous genus Erythrobasidium as Erythrobasidium hasegawae comb. nov. Journal of General and Applied Microbiology 34: 279-287.
Hamamoto, M., Thanh, V.N. and Nakase, T. 2002. Bannoa hahajimensis gen. nov., sp. nov., and three related anamorphs, Sporobolomyces bischofiae sp. nov., Sporobolomyces ogasawarensis sp. nov. and Sporobolomyces syzygii sp. nov., yeasts isolated from plants in Japan. International Journal of Systematic and Evolutionary Microbiology 52: 1023-1032.
Kirschner, R., Sampaio, J.P., Gadanho, M., Weiss, M. and Oberwinkler, F. 2001. Cuniculitrema polymorpha (Tremellales, gen. nov. and sp. nov.), a heterobasidiomycete vectored by bark beetles, which is the teleomorph of Sterigmatosporidium polymorphum. Antonie van Leeuwenhoek 80: 149-161.
McLaughlin, D.J. 1990. A new cytoplasmic structure in the basidiomycete Helicogloea: the microscala. Experimental Mycology 14: 331-338.
McLaughlin, D.J., Berres, M.E. and Szabo, L.J. 1995a. Molecules and morphology in basidiomycete phylogeny. Canadian Journal of Botany 73: 684-692.
McLaughlin, D.J., Frieders, E.M. and Lü, H. 1995b. A microscopist’s view of heterobasidiomycete phylogeny. Studies in Mycology 38: 91-109.
Oberwinkler, Fand Bauer, R. 1989. The systematics of gasteroid, auricularioid Heterobasidiomycetes. Sydowia 41: 224-256.
Prillinger, H., Dörfler, Ch., Laaser, G. e Hauska, G. 1990. Ein Beitrag zur Systematik und Entwicklungsbiologie Höherer Pilze. Hefe-Typen der Basidiomyceten. Teil III:Ustilago-Typ. Zeitschrift für Mykologie 56: 251-278.Prillinger, H., Laaser, G., Dörfler, Ch., e Ziegler, K. 1991a. Ein Beitrag zur Systematik und Entwicklungsbiologie Höherer Pilze. Hefe-Typen der Basidiomyceten. Teil IV: Dacrymyces-Typ, Tremella-Typ. Sydowia 53: 170-218.
Prillinger, H., Deml, G., Dörfler, Ch., Laaser, G. e Lockau, W. 1991b. Ein Beitrag zur Systematik und Entwicklungsbiologie Höherer Pilze. Hefe-Typen der Basidiomyceten. Teil II: Microbotryum-Typ. Botanica Acta 104: 5-17.
Prillinger, H., Oberwinkler, F., Umile, C., Tlachac, K., Bauer, R., Dörfler, C. e Taufratzhofer, E. 1993. Analysis of cell wall carbohydrates (neutral sugars) from ascomycetous and basidiomycetous yeasts with and without derivatization. Journal of General and Applied Microbiology 39: 1-34.
Sampaio, J.P., Bauer, R., Begerow, D. andOberwinkler, F. 1999. Occultifur externus sp. nov., a new species of simple-pored auricularioid heterobasidiomycete from plant litter in Portugal. Mycologia 91: 1094-1101.
Sampaio, J.P., Weiß, M., Gadanho, M. and Bauer, R. 2002. New taxa in the Tremellales: Bulleribasidium oberjochense gen. et sp. nov., Papiliotrema bandonii gen. et sp. nov. and Fibulobasidium murrhardtense sp. nov. Mycologia 94: 873-887.
Sampaio, J.P., Gadanho, M., Bauer, R. and Weiß, M. 2003. Taxonomic studies in the Microbotryomycetidae: Leucosporidium golubevii sp. nov., Leucosporidiella gen. nov. and the new orders Leucosporidiales and Sporidiobolales. Mycological Progress (in press).
Scorzetti, G., Fell, J.W., Fonseca, A. and Statzell-Tallman, A. 2002. Systematics of basidiomycetous yeasts: a comparison of large subunit D1/D2 and internal transcribed spacer rDNA regions. FEMS Yeast Research 2: 495-517.
Swann, E.C. and Taylor, J.W. 1993. Higher taxa of basidiomycetes: an 18S rRNA gene perspective. Mycologia 85: 923-936.
Swann, E.C. and Taylor, J.W. 1995a. Phylogenetic perspectives on basidiomycete systematics: evidence from the 18S rRNA gene. Canadian Journal of Botany 73: 649-1433.
Swann, E.C. and Taylor, J.W. 1995b. Phylogenetic diversity of yeast-producing basidiomycetes. Mycological Research 99: 1205-1210.
Swann, E.C. and Taylor, J.W. 1995c. Toward a phylogenetic systematics of the Basidiomycota: integrating yeasts and filamentous basidiomycetes using 18S rRNA gene sequences. Studies in Mycology 38: 147-161.
Swann, E.C. Frieders, E.M. and McLaughlin, D.J. 1999. Microbotryum, Kriegeria and the changing paradigm in basidiomycete classification. Mycologia 91: 51-66.Swann, E.C. Frieders, E.M. and McLaughlin, D.J. 2001. Urediniomycetes In The Mycota – vol VII, Systematics and Evolution, D.J. McLaughlin, E.G. McLaughlin and P.A. Lemke (eds.), Springer-Verlag, Berlin, pp 37-56
Takashima, M., Hamamoto, M. and Nakase, T. 2000. Taxonomic significance of fucose in the class Urediniomycetes: distribution of fucose in cell wall and phylogeny of urediniomycetous yeasts. Systematic and Applied Microbiology 23: 63-70.
Taylor, J.W. 1995. Molecular phylogenetic classification of fungi. Archives of Medical Research 26: 307-314.
Wells, K. 1994. Jelly fungi, then and now! Mycologia 86: 18-48.
Wells, K. and Bandoni, R.J. 2001. Heterobasidiomycetes. In The Mycota – vol VII, Systematics and Evolution, D.J. McLaughlin, E.G. McLaughlin and P.A. Lemke (eds.), Springer-Verlag, Berlin, pp 85-120.
![]() About this page
About this page
Sampaio, J.P. and Bauer, R. 2003. The classification of dimorphic basidiomycetes. Dimorphic Basidiomycetes WWW Project.
Version 01, saved 25 March 2003.
José Paulo Sampaio: jss@fct.unl.pt
Centro de Recursos Microbiológicos, Secção Autónoma de Biotecnologia, Faculdade de Ciências e Tecnologia, Universidade Nova de Lisboa, 2829-516 Caparica, Portugal.
Robert Bauer: robert.bauer@uni-tuebingen.de
Universität Tübingen, Institut für Biologie I, Lehrstuhl Spezielle Botanik und Mykologie, Auf der Morgenstelle 1, D-72076 Tübingen, Germany
Page copyright © 2003 J.P. Sampaio and R. Bauer
Dimorphic Basidiomycetes design and icons copyright © 2002 J.P. Sampaio